

A brief history of the human population on Earth
A story in four parts

[I will be teaching a class on human fertility decline at the University of Chicago in the Spring 2025 quarter, beginning in April. I am giving 8-10 lectures. One of the first will address the history of human population (levels and growth). This essay covers some of the topics I will address in that lecture. In the segment below, I cover only the first two of the four parts of the history of human population. I plan to post an essay or two on the other parts in the next week or so. For those who are reading, the references I cite are a reasonable reading list for any individuals interested in dividing deeper into the history of human population.]
The history of human–i.e., Homo sapien—population can be divided into four parts. Being present- and policy-focused, we are tempted to spend most of our time on the fourth, most modern part. However, prior segments provide useful lessons about the biggest drivers of population growth.
The first part is the period before homo sapiens emerge. This includes the emergence of the genus homo from apes, specifically chimpanzees or bonobos, and the proliferation of non-sapien homo species. The second period between the birth of sapiens and the Neolithic Revolution. This period includes our emergence from Africa and focuses on hunting and gathering for sustenance. The third part spans the period between Neolithic and Industrial Revolutions. It is the age of agriculture, growth in settlements and empires, and technological advances that precursor the rapidity of the last period. The fourth period takes us from the Industrial Revolution to the present. It is the age of manufacturing, but includes much else. It encompasses the Demographic Transition, wherein the decline in mortality rates led to a dramatic growth in population. But this growth was followed by even larger, continued declines in fertility, that imply an imminent (in a historical sense) peak of human population on Earth.
In this essay I unpack the first two periods.
A framework
Before I begin, I want to provide a framework to think about human population size that will help us understand the drivers of population growth in each human era. Human population at any given time is
Human population = (humans/sq. km) x (sq km. occupied by humans) = density x area
In my description of human history, you will see that early human population growth is driven by their geographic spread, i.e., the area they occupy. Later human growth is driven by the density of their population. When examining the role that technology and social organization plays in human population growth, we see this same pattern. Early technology and organization favors expansion of area, while later developments favor density.
Of course, this framework is a simplification. Early humans may have spread around the globe before the Neolithic Revolution, but they did not occupy all ecological niches between them. So the agricultural revolution involved both finding new niches and increasing density in each niche. Moreover, development after the Industrial Revolution involved substantial urbanization. As agriculture became more efficient and capital intensive, humans moved from rural areas to cities. The number of niches they occupied remained the same, but they were not evenly distributed across those niches. This is made possible by trade of agricultural goods for other goods. Nevertheless, the area versus density framework is useful for understanding why human population grew across the millenia.
1. The before times (8 mya - 300 kya = ~8 my)
Humans, i.e., Homo sapiens, are a species of the genus Homo. Chimpanzees and bonobos are members of genus Pan. The two genera, Homo and Pan, are sister clades that share a common ancestor (de Manuel et al., 2016).1 The human lineage and the lineage that led to chimps and bonobos took place 6-8 mya (million years ago) (Patterson et al., 2006). The modern homo genus emerged roughly 2.5-2.8 mya, though sapiens emerged much later (Antón, 2012; Villmoare et al., 2015).
There are several features that distinguish the genus Homo genus from its sister clade (Tattersall, 2012). One is large brain size, which enables advanced cognitive functioning such as abstract reasoning, planning and problem solving (Lieberman, 2011). Another is the use of more sophisticated tools to augment its physical capacity, allowing Homo to exploit new food sources (Ambrose, 2001). A third is symbolic thought and complex language (McBrearty & Brooks, 2000). This enables the ability to create culture and transmit knowledge across a living population and generations. (Arguably brain size is responsible for sophisticated tool use and symbolic thought and language, the main feature is brain size.) When combined with bipedalism (Bramble & Lieberman, 2004; Carrier et al., 1984), which allowed more calorie-efficient mobility, these features enabled Homo to colonize a wider range of habitats—from the tropics to the Arctic. Indeed, the geographic range of Homo surpasses nearly all, if not all, large-bodied land animals.

Source: https://en.m.wikipedia.org/wiki/File:Pan_%28genus%29_distribution_map.svg
{kind=link}
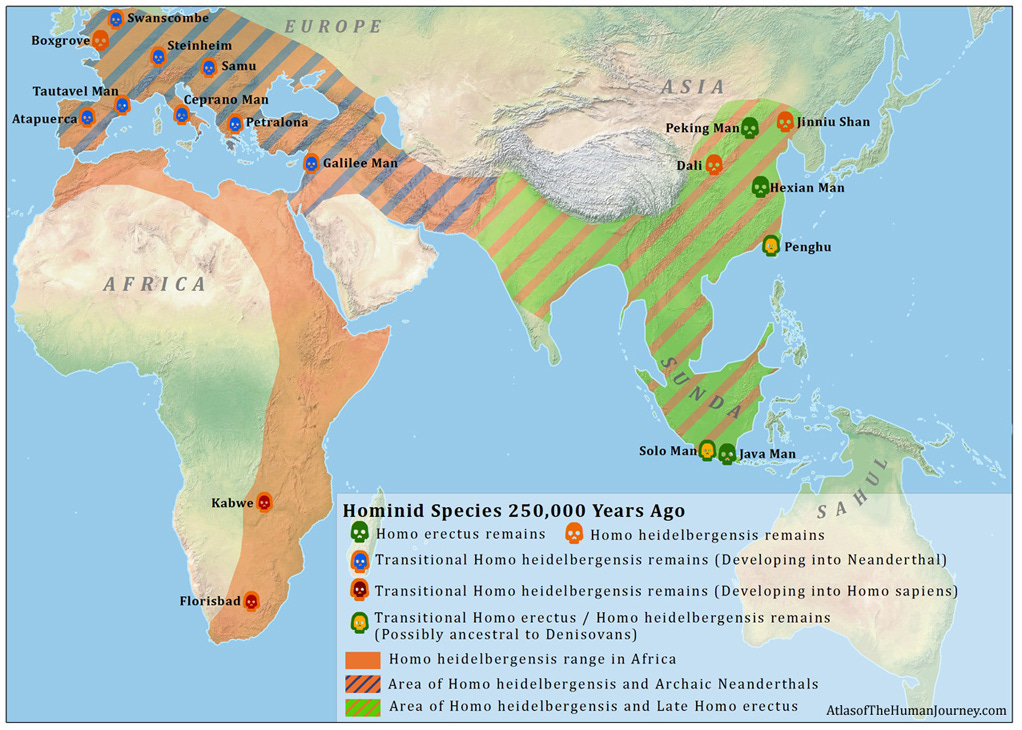
Source: http://atlasofhumanevolution.com/HomoErectus.asp
The maps above show the geographic scope of Pan and Homo (excluding sapiens). This range is not the same as population level. It is difficult to calculate the population of Pan, as they do not bury their dead (preserving skeletons) or live in areas conducive to fossil preservations. We have more information on Homo specimens. But even this is difficult because not all Homo species bury their dead. The important lesson for our purposes is that, until sapiens emerged roughly 200-300 kya (thousand years ago), there were no modern humans.
2. The Paleolithic Era (300 kya - 10 kya = ~300 kya)
Humans were not the only Homo species on Earth during the Paleolithic era. Others include H. naledi, H. heidelbergensis, and H. rudolfensis in Africa, Neanderthals and Denisovians in Eurasia, and H. flouriensis, H. luzonensis and late archaic humans in Asia, among others. At the start of the Paleolithic period, humans were a small fraction of Homo species on Earth, appearing only in Africa. By 30 kya, all the other Homo species had disappeared. Humans interbred with Neanderthals and Denisovians, so the latter’s genes survive in humans today. But sapiens not only colonized all the areas occupied by other Homo species, but many regions beyond.

Source: https://sociology.morrisville.edu/Class%20Notes/ANTH101/-%20Exam%20Review%20Sheets/18SP-anthro-review-exam1.html
There are several reasons humans survived other Homo species. They arguably had greater capacity for abstract thought (Bouzouggar et al., 2007). Sapiens may have greater capacity to make sophisticated tools and technology (Shipman, 2015). Some have argued that sapiens were better adapted to a warming world, facilitating their spread (Shichi et al.). Finally, humans may have had lower child mortality, allowing them to have a higher effective fertility rate (Dennell & Hurcombe, 2024). Importantly, these advantages were sufficiently large that they outweighed the limitations of the species: its small initial size and reduced physical robustness relative to, e.g., Neanderthals, who were better adapted to extreme cold without reliance on technology (Pomeroy, 2023).
Out of Africa
Homo sapiens originated in Africa, the same continent that the earliest Homo species (habilis) originated, roughly 200-300 kya. Although there were several waves of humans that emerged from Africa, the first wave that survived outside Africa emigrated from that continent roughly 60 kya (Henn et al., 2012; Mellars, 2006).
We know there were no humans just before sapiens evolved. But how many were there when humans successfully ventured out of Africa 60 kya? The answer depends on how you define “population”.
The global census population estimates how many humans there actually were on Earth. That number is quite low, perhaps just 10,000-50,000 persons. (By contrast, it is estimated there are 170,000-300,000 chimps alive today. See https://www.ifaw.org/animals/chimpanzees#368138.) Moreover, archaeological evidence suggests humans were confined to a limited area of Africa. Both these facts reinforce the idea that our existence was quite precarious relatively recently in our planet’s history.
While the global population census estimate is quite small, it is not as small as the effective population size, which describes the number of breeding individuals in a hypothetical population that would experience the same genetic drift as observed in the actual population. The limited genetic variability in modern humans suggests the effective population size was just 1,000-2,000 when humans first left Africa (Henn et al., 2012). This suggests a serious genetic bottleneck in our past. It also explains why humans are thought to have limited genetic variability as compared to chimpanzees, even though our modern global population census exceeds 8 billion souls, 4 orders of magnitude larger than the population of chimps.
After emerging from Africa, however, humans spread further across the globe than any other Homo species. By the Neolithic Revolution, they may have occupied every continent save Antarctica, and spanned the Arctic to the southernmost tip of South America.

Source: http://atlasofhumanevolution.com/HomoSapiens.asp
Moreover, the human population expanded substantially after expanding out of Africa. After growing, by just 50 thousand persons over 200,000 years in Africa, the human population grew to 1-10 million persons by 10 kya (Bocquet-Appel, 2011). If we assume that humans grew from a population of 2 in 250 kya to a population of 50,000 in 60 kya, then the human population growth rate was ~0.0053% per year (0.5%/century) over that roughly 200,000 year span. If we assume that humans grew from a population of 50,000 to a population of 5 million from 60 kya to 10 kya, then the human population grew by ~0.0092% per year (~1%/century), over that 50,000 year span, double the rate before emerging from Africa. Average growth during the entire period from the appearance of sapiens on Earth to the Neolithic Revolution is ~0.0061% per year.
Hunting and gathering
Whereas plants consume sunlight, carbon dioxide and water and use photosynthesis to convert that to chemical energy (ATP) stored in carbohydrates (predominantly glucose) and lipids (fat), animals consume other organic matter, oxygen and water and use enzymatic processes, often in combination with gut bacteria, to convert that into chemical energy (ATP) stored as carbohydrates and lipids (Nelson et al., 2021; Urry et al., 2021). A challenge for all animals is obtaining these inputs, though for land based animals, the primary challenge is obtaining fresh water and organic matter.2
Although nearly all animals engage in some form of hunting (predation and scavenging3) and/or gathering (foraging), the term hunting and gathering is uniquely associated with the genus Homo.4 There is no qualitative distinction between the behavior of Homo and other genera (including non-Homo primates). Rather the differences in degree of sophistication in these activities, notably the use of tools and technological processes to obtain organic matter (Boesch, 1994; DeVore et al., 1968; Wrangham, 2009).
Hunting and gathering among Homo species evolved during the Paleolithic era (Ambrose, 2001). Early species, such as habilis, used basic stone tools, was opportunistic (i.e., involved little planning), and engaged in scavenging. By the middle paleolithic period, Homo developed more complex tools and engaged in group pursuit of game, an early form of social organization. By the later Paleolithic period, sapiens introduced a wide range of specialized tools, more sophisticated hunting strategies, and developed an ability to obtain food in diverse ecosystems, from coasts to inland forests (i.e., developed and transmitted ecological knowledge).
A few technological developments played critical roles in improving hunting and gathering, allowing humans to both spread geographically and to increase in population. One is fire, which was first used in a controlled fashion by Homo erectus roughly 1 mya (Wrangham, 2009). Fire enabled cooking, which increased digestibility and safety of food. It also enabled survival in cold climates. A second is advanced tools, such as axes that enabled butchering animals and spears (but not hooks) for fishing (Ambrose, 2001; Wrangham et al., 1999). A third is clothing (~ 70-170 kya) and shelter (~40 kya), which enabled survival in colder environments (Ambrose, 2001; Henshilwood & Marean, 2003; Toups et al., 2011).
Two important features of modern human societies emerged during the later Paleolithic Era, especially among sapiens. One is the development of villages. Whereas other Homo species occasionally used temporary camps, sapiens began permanent villages in the Upper Paleolithic period (~10-40 kya). The other is trade, i.e., exchange of different goods in a manner that benefited both parties to the trade. While Neanderthals appear to have engaged in some trade, Upper Paleolithic sapiens show more systematic, long-distance exchange of materials. The latter demonstrate a more sophisticated trade network involving larger social groups (Mellars, 2006).
Neither agglomeration or trade is critical to hunting and gathering. Even Upper Paleolithic humans have fairly rudimentary settlements and exchange. But these two characteristics of human social organization take off after the Neolithic Revolution introduces a new technology for producing food: agriculture. Moreover, cities and trade, both of which enable specialization and innovation, play a central role in helping humans escape the harsh tradeoffs of Malthusian economics and raise their population growth rate after the Industrial Revolution.
Acknowledgements
I think Stacy Rosenbaum for comments on this draft.
References
Ambrose, S. H. (2001). Paleolithic Technology and Human Evolution. Science, 291(5509), 1748-1753. https://doi.org/10.1126/science.1059487
Antón, S. C. (2012). Early Homo: Who, When, and Where. Current Anthropology, 53(S6), S278-S298. https://doi.org/10.1086/667695
Bocquet-Appel, J.-P. (2011). When the World’s Population Took Off: The Springboard of the Neolithic Demographic Transition. Science, 333(6042), 560-561. https://doi.org/10.1126/science.1208880
Boesch, C. (1994). Cooperative hunting in wild chimpanzees. Animal Behaviour, 48(3), 653-667. https://doi.org/https://doi.org/10.1006/anbe.1994.1285
Bouzouggar, A., Barton, N., Vanhaeren, M., d'Errico, F., Collcutt, S., Higham, T., Hodge, E., Parfitt, S., Rhodes, E., Schwenninger, J.-L., Stringer, C., Turner, E., Ward, S., Moutmir, A., & Stambouli, A. (2007). 82,000-year-old shell beads from North Africa and implications for the origins of modern human behavior. Proceedings of the National Academy of Sciences, 104(24), 9964-9969. https://doi.org/10.1073/pnas.0703877104
Bramble, D. M., & Lieberman, D. E. (2004). Endurance running and the evolution of Homo. Nature, 432(7015), 345-352. https://doi.org/10.1038/nature03052
Carrier, D. R., Kapoor, A. K., Kimura, T., Nickels, M. K., Scott, E. C., So, J. K., & Trinkaus, E. (1984). The Energetic Paradox of Human Running and Hominid Evolution [and Comments and Reply]. Current Anthropology, 25(4), 483-495. https://doi.org/10.1086/203165
de Manuel, M., Kuhlwilm, M., Frandsen, P., Sousa, V. C., Desai, T., Prado-Martinez, J., Hernandez-Rodriguez, J., Dupanloup, I., Lao, O., Hallast, P., Schmidt, J. M., Heredia-Genestar, J. M., Benazzo, A., Barbujani, G., Peter, B. M., Kuderna, L. F. K., Casals, F., Angedakin, S., Arandjelovic, M.,…Marques-Bonet, T. (2016). Chimpanzee genomic diversity reveals ancient admixture with bonobos. Science, 354(6311), 477-481. https://doi.org/10.1126/science.aag2602
Dennell, R., & Hurcombe, L. (2024). How and why is Homo sapiens so successful? Quaternary Environments and Humans, 2(2), 100006. https://doi.org/https://doi.org/10.1016/j.qeh.2024.100006
DeVore, I., Lee, R. B., & De Vore, I. (1968). Man the hunter. Aldine Publishing Company Chicago, IL.
Henn, B. M., Cavalli-Sforza, L. L., & Feldman, M. W. (2012). The great human expansion. Proceedings of the National Academy of Sciences, 109(44), 17758-17764. https://doi.org/10.1073/pnas.1212380109
Henshilwood, Christopher S., & Marean, Curtis W. (2003). The Origin of Modern Human Behavior: Critique of the Models and Their Test Implications. Current Anthropology, 44(5), 627-651. https://doi.org/10.1086/377665
Lieberman, D. E. (2011). The Evolution of the Human Head. Harvard University Press. https://doi.org/doi:10.4159/9780674059443
McBrearty, S., & Brooks, A. S. (2000). The revolution that wasn't: a new interpretation of the origin of modern human behavior. Journal of Human Evolution, 39(5), 453-563. https://doi.org/https://doi.org/10.1006/jhev.2000.0435
Mellars, P. (2006). Why did modern human populations disperse from Africa ca. 60,000 years ago? A new model. Proceedings of the National Academy of Sciences, 103(25), 9381-9386. https://doi.org/10.1073/pnas.0510792103
Nelson, D. L., Cox, M. M., & Hoskins, A. A. (2021). Lehninger Principles of Biochemistry. Macmillan Learning. https://books.google.com/books?id=5faXzQEACAAJ
Patterson, N., Richter, D. J., Gnerre, S., Lander, E. S., & Reich, D. (2006). Genetic evidence for complex speciation of humans and chimpanzees. Nature, 441(7097), 1103-1108. https://doi.org/10.1038/nature04789
Pomeroy, E. (2023). Review: The different adaptive trajectories in Neanderthals and Homo sapiens and their implications for contemporary human physiological variation. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 280, 111420. https://doi.org/https://doi.org/10.1016/j.cbpa.2023.111420
Shichi, K., Goebel, T., Izuho, M., & Kashiwaya, K. Climate amelioration, abrupt vegetation recovery, and the dispersal of Homo sapiens in Baikal Siberia. Science Advances, 9(38), eadi0189. https://doi.org/10.1126/sciadv.adi0189
Shipman, P. (2015). The Invaders. Harvard University Press. https://doi.org/doi:10.4159/9780674425385
Tattersall, I. (2012). Masters of the Planet: The Search for Our Human Origins. St. Martin's Publishing Group. https://books.google.com/books?id=h5PGjJW8FLoC
Toups, M. A., Kitchen, A., Light, J. E., & Reed, D. L. (2011). Origin of Clothing Lice Indicates Early Clothing Use by Anatomically Modern Humans in Africa. Mol Biol Evol, 28(1), 29-32. https://doi.org/10.1093/molbev/msq234
Urry, L. A., Cain, M. L., Wasserman, S. A., Minorsky, P. V., & Orr, R. B. (2021). Campbell Biology. Pearson. https://books.google.com/books?id=zqFlxwEACAAJ
Villmoare, B., Kimbel, W. H., Seyoum, C., Campisano, C. J., DiMaggio, E. N., Rowan, J., Braun, D. R., Arrowsmith, J. R., & Reed, K. E. (2015). Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia. Science, 347(6228), 1352-1355. https://doi.org/10.1126/science.aaa1343
Wrangham, R. (2009). Catching fire: how cooking made us human. Basic books.
Wrangham, Richard W., Jones, James H., Laden, G., Pilbeam, D., & Conklin‐Brittain, N. (1999). The Raw and the Stolen: Cooking and the Ecology of Human Origins. Current Anthropology, 40(5), 567-594. https://doi.org/10.1086/300083
Between Pan and Homo is a genus called Australopithecus, believed to be an ancestor of Homo. Hominini split into Pan and Hominina, Australopithecus descended from Hominina, and Homo descended from Australopithecus. See https://en.wikipedia.org/wiki/Hominini.
Fish face the challenge of obtaining oxygen and organic matter, as they swim in water. That said, saltwater fish need to use a process called osmoregulation to filter out salt in saltwater.
Predation involves killing a live animal. Scavenging is the consumption of dead animals.
Some animals, such as bivalves, some fish, and sponges engage in filter feeding. Parasites extract nutrients from a host. These activities are neither predation, scavenging or foraging.